Весенние источники инфекции и прогноз первичных заражений
В условиях Северо-Запада роль первичной весенней инфекции принадлежит аскоспорам гриба. При специальных маршрутных обследованиях садов нами не обнаружены какие-либо другие источники весенней инфекции — ни зимующие конидии, ни мицелий гриба. Также не обнаружено поражения ветвей яблони грибом. Тем не менее в СССР есть местности, где сумчатая стадия гриба в отдельные годы может не образовываться. Например, в Средней Азии гриб способен перезимовывать конидиями и мицелием на пораженных ветвях и листьях яблони и служить дополнительным источником инфекции, причем поражает гриб в основном дикие яблони.
В условиях Сибири, по данным профессора Томского университета Н. Н. Лаврова (по Э. Э. Гешеле, 1958), распространение парши яблони и развитие паразита возможно только в конидиальной стадии, и гриб в данном случае поражает не только листья и плоды, но и побеги, завязи и плодоножки. Однако Э. Э. Гешеле утверждает, что возбудитель парши яблони имеет в Сибири и сумчатую стадию, что было впервые указано для Омской области в 1946 г. Процесс заражения яблони сумкоспорами может происходить здесь с мая по сентябрь, но массовое заражение происходит во второй половине июля, т. е. в период июльского максимума осадков. В Сибири аскоспоры созревают очень неравномерно. Например, на 14 июля 1948 г. приходилось 7,2% сумок с незрелыми сумкоспорами, на 20 августа 1950 г. — 10% и в сентябре того же года было 8,5%.
По данным А. Ермасовой (1906), в районе г. Сызрани и соседних приволжских селах сумчатая стадия также не наблюдается. Поэтому было необходимо выяснить ряд вопросов, чтобы иметь достоверное суждение о фактическом ходе заражения и нужных для этого условиях.
Способы распространения и дальность рассеивания аскоспор. У грибов наблюдаются самые различные механизмы и способы рассеивания и распространения спор. Споры грибов обладают различными приспособлениями для воздушного распространения. Например, у аскомицетов наблюдается баллистический механизм выбрасывания опор из сумок; его называют водяной пушкой (Ф. Грегори, 1954). Ц. Ингольд (1957) установила у аскомицетов 4 четко очерченных типа освобождения опор. К одному из них относится изучаемый гриб V. inaequalis. Этот тип освобождения характерен для пиреномицетов. У них сумки находятся в небольшом кувшиновидном органе — перитеции, который открывается наружу маленькой порой (устьицем). Для того чтобы сумка могла выбросить споры, ее конец должен достигнуть устьица, а так как канал шейки перитеция очень узок, сумки могут выходить только поочередно.
Проблема воздушного распространения опор грибов привлекла большое внимание ученых. В различных странах была создана специальная аппаратура для взятия проб воздуха различными методами. Исследование проб воздуха привело к обнаружению разнообразия организмов, распространяющихся воздушными спорами. Это помогло установить условия распространения эпифитотий и изменение концентрации воздушных спор в зависимости от их удаления от источника.
Явление суточных колебаний концентраций спор детально изучал J. M. Hirst (по Ф. Грегори, 1964). Им установлены некоторые характерные типы суточной периодичности и выделены «ночной», «утренний», «дневной» и «вечерний» типы. Аскоспоры сумчатых относятся им к «ночному» типу, характеризующемуся наивысшей концентрацией спор в воздухе.
Время года оказывает еще большее влияние на концентрацию воздушных спор, чем время суток. Споры V. inaeqyalis приурочены в основном к весенне-летнему периоду.
Е. D. Hamilton (по Ф. Грегори, 1964) приводит данные о температуре, при которой различные типы спор присутствуют в воздухе в максимальной концентрации. Для спор Vcnturia эта температура лежит в пределах от 7,2 до 9,4°С, а для опор Ustilago для сравнения — от 21,1 до 23,3°С.
Какова же дальность рассеивания аскоспор V. inaequalis? Обычно аскоспоры выбрасываются из перитециев на высоту 30 — 50 мм, подхватываются воздушными токами и разносятся у поверхности земли. В опытах, проводимых нами по улавливанию аскоспор из воздуха, которые будут подробно описаны далее, ловушки выставлялись как на поверхности почвы, так и на расстоянии одного и 2 м от поверхности земли. В опытах по улавливанию спор ловушками, расположенными непосредственно на земле, спор было всегда несколько больше, нежели на высоте 2 м; видимо, в этом случае выброшенные из перитециев опоры, не подхваченные токами воздуха, сразу же оседали в ловушке.
Наличие микроорганизмов в верхних слоях атмосферы одним из первых установил миколог С. O. Harz. Он проводил забор проб воздуха через фильтр, установленный на аэростате на высоте от 1500 до 2300 м. Количество микроорганизмов в верхних слоях атмосферы невелико, но все же достаточно, и к тому же они жизнеспособны, т. е. после улавливания хорошо прорастают в культуре.
Споры грибов из рода Venturia были обнаружены во время перелета из США (штат Мэн) в Данию Линдбергом, который экспонировал на высоте 1000 м клейкие предметные стекла, установленные в специальных контейнерах.
Микроорганизмы обнаружены в воздухе даже на полюсах Земли, хотя распространение их там очень неравномерно.
Сроки рассеивания аскоспор. Сроки рассеивания аскоспор в очень большой степени зависят от метеорологических условий данной местности, а также от сроков образования и созревания перитециев, что в свою очередь зависит от времени листопада, сорта яблонь, температуры и влажности осенних месяцев, в которые при благоприятных условиях может происходить закладка перитециев, и от зимних и весенних температур и осадков.
В нашей стране наблюдения за развитием сумок и сроками лёта аскоспор ведутся в основном визуальным методом.
Созревание сумок в разных климатических зонах страны и даже в одном и том же районе происходит в разное время. Так, по наблюдениям В. Я. Франковского (1952), в Тернопольской области в 1949 г. окрашенные сумкоспоры в перитециях наблюдались в течение марта, а в 1950 г. — лишь 5 апреля. Созревание происходит в зависимости от погоды осенью и весной. Теплые осень и весна с осадками обусловливают более быстрое созревание спор. В Харьковской области созревание аскоспор в перитециях было отмечено: в 1965 г. — 26 апреля, в 1966 г. — 7 августа, в 1967 г. — 4 апреля (Г. В. Литвинова, 1969) и т. д.
Опыты по установлению сроков первичной инфекции и продолжительности лёта аскоспор автором проводились в учебно-опытном хозяйстве Ленинградского сельскохозяйственного института в течение 1964—1966 гг. Аскоспоры улавливались гравитационным методом (естественное оседание на агаризированные чашки Петри) и учитывались количественно на 1 см2 поверхности.
Благодаря этим опытам удалось установить начало лёта аскоспор: в 1964 г. лёт аскоспор начался 2 мая после выпадения 1,6 мм осадков (яблоневые почки в это время еще не пробудились) и продолжался до конца вегетационного периода.
В 1965 г. лёт аскоспор начался 11 мая после выпадения 2,2 мм осадков в фазе выдвижения соцветий у ранних сортов и продолжался до августа.
В 1966 г. лёт аскоспор начался 9 мая в фазе зеленого конуса после выпадения 7 мая 2,1 мм, 8 мая — 4 мм осадков.
Как видим, лёт аскоспор происходит в различные фазы развития яблонь.
В 30-х годах как в зарубежной, так и в отечественной литературе, часто рекомендовали опрыскивания по фенофазам развития яблонь. Причиной этому послужили наблюдавшиеся случаи совпадения максимального рассеивания аскоспор с определенной фенофазой яблони. На Украине с 1934 по 1951 г. наиболее распространенной схемой опрыскивания была схема О. Н. Югановой.
Кратковременный прогноз первичных заражений яблони паршой. Очень важно заранее предвидеть начало отдельных заражений растений и проявления болезней, а также степень ожидаемого их развития. Благодаря этому можно заранее устанавливать объем и сроки проведения борьбы. Чтобы поставить прогноз той или иной болезни, надо учитывать запас инфекционного начала, устойчивость сортов и состояние факторов среды, главным образом погоды, так как именно погодные факторы имеют решающее значение.
Различают две формы сезонного прогноза: краткосрочный и долгосрочный. Краткосрочный прогноз позволяет предвидеть сроки отдельных заражений и проявлений болезней и имеет значение для сигнализации защитных опрыскиваний или опыливаний.
Долгосрочный позволяет предвидеть за продолжительный период времени вероятный размер развития болезни.
Многие исследователи сходятся на том, что численность аскоспор в воздухе находится в прямой зависимости от количества весенних осадков. Однако в литературе по этому вопросу нет подтверждения обнаруженной связи и не приводятся количественные показатели существующей корреляции, пригодные для прогноза.
Для установления такой связи нами был проведен следующий опыт.
Учитывались количество осевших аcкоспор на 1 см2 поверхности (у), суммарная продолжительность периодов выпадения осадков в минутах за сутки до учета (x) и сумма осадков в миллиметрах в тот же срок (z). После математической обработки данных выяснилось, что связь количества аcкоспор с продолжительностью дождей достаточно тесная (rух — 0,71), с суммой осадков — средняя (rуz = 0,53), а количества выпавших осадков с их продолжительностью очень тесная (rxz = 0,85). Исследование последнего обстоятельства с помощью парциальных (частных) коэффициентов корреляции показало, что связь количества аокоспор с продолжительностью дождя при исключении влияния суммы осадков почти не изменяется (ryx•z=0,7), связь же количества аскоспор с суммой осадков при исключении влияния продолжительности дождей оказалась равной нулю (ryz•x = 0). Это математическое положение можно подтвердить и биологически.
Допустим, прошел сильный ливень и выпало большое количество осадков, однако дождь шел недолго, и перитеции, начав освобождаться от аскоспор, прекратили рассеивать их из-за того, что листья стали подсыхать на солнце и ветру, не получая больше влаги. В другом случае — количество осадков при моросящем дожде бывает небольшим, однако листья все время находятся во влажном состоянии, и лёт аскоспор происходит в продолжение дождя.
Таким образом, нами было впервые доказано, что количество аскоспор в воздухе зависит главным образом от продолжительности увлажнения подстилки (перезимовавших листьев), а не от количества выпавших осадков. На этом основании было выведено уравнение регрессии, выражающее интересующую нас связь в аналитической форме: r=5,4х + 446.
Зная продолжительность выпавших дождей в минутах, можно вычислить ожидаемую степень заражения поверхности листьев (число аскоспор на 1 см2) на следующий день после дождя, т. е. прогнозировать интенсивность лёта аскоспор гриба.
Чтобы составить прогноз интенсивности лёта аcкоспор и определить необходимость опрыскивания против парши яблони, удобнее пользоваться шкалой, составленной на основании приведенного выше уравнения и имеющей производственное значение.

Таблица 1. Шкала для оценки заспоренности поверхности аскоспорами в зависимости от продолжительности осадков
Влияние влажности и температуры на процесс заражения яблонь паршой. Наличие определенной влажности оказывает большое влияние на прорастание спор и начальное развитие большинства возбудителей болезней, споры которых оседают на растения из воздуха. У этих возбудителей максимальная влажность является и оптимальной, поэтому заражение происходит в капле воды, т. е. во время росы, тумана или дождя. К таким аэрогенным возбудителям относится и гриб V. inaequalis, вызывающий паршу яблонь.
Споры могут прорастать и во влажном воздухе, так как вследствие переохлаждения тканей листа в результате транспирации уже при 98% относительной влажности воздуха вокруг самих спор и ростковых трубок образуется, очевидно, тонкий слой (конденсированной влаги (Э. Гойман, 1954). Однако с понижением влажности воздуха прорастание спор все же резко уменьшается.
Таким образом, необходимость очень высокой влажности в процессе заражения яблонь паршой неоспорима и не вызывает сомнений. Поэтому наши исследования были направлены на изучение влияния температуры окружающей среды на процесс заражения.
Для заражения растений значение температуры окружающей среды несколько меньше, чем влажности, так как температура только активизирует или замедляет процесс заражения. Но температура окружающей среды влияет на образование опор, на скорость их прорастания и на скорость роста ростковых трубок.
Литературные данные о влиянии температуры на прорастание аскоспор гриба V. inaequalis несколько разноречивы. Так, у О. Н. Югановой (1934) оптимальной температурой является 15—20°С, В. Я. Франковский (1952) указывает минимальную температуру при прорастании аскоспор 3°С, оптимальную 19 — 25°С, максимальную 30°С. По А. Л. Ячевскому (1933), минимальная температура 6°С, оптимальная 20—22°С, максимальная 33°С; по R. Aderhold (1896), — минимальная 11—12°С, оптимальная 22°С; по G. W. Keitt, L. К. Jones (1926), минимальная 2°С, оптимальная 22°С, максимальная 33°С.
Нами были проведены исследования по проращиванию аскоспор гриба, взятых с перезимовавших листьев яблонь сорта Пепин-китайка.
Для этой цели приготавливалась аcкоспоровая суспензия: кусочки перезимовавших листьев с перитециями помещались во влажную камеру — кольцо Ван-Тигема, которое накрывалось покровным стеклом с висячей каплей дистиллированной воды. Аскоспоры выбрасывались из перитециев и попадали в эту каплю. Влажные камеры затем помещались на 24 ч в политермостат с температурой в камерах 2, 6, 8, 14, 22, 27, 32°С.
Опыт показал, что оптимальные температуры для прорастания аскоспор северо-западной популяции гриба лежат в пределах от 14 до 27°С, но 22°С следует считать наиболее благоприятной, так как при этом наблюдается не только 100%-ное прорастание спор, но и наличие большого количества спор, проросших двумя ростками, т. е. каждая клетка аскоспоры дала росток. При этой температуре наблюдается также и наибольшая длина ростковых трубок. Начало прорастания отмечается при 8°С; замедляется прорастание при 32°С.
Влияние температуры на длительность инкубационного периода при заражении аскоспорами. Длина инкубационного периода зависит от внешних условий (температуры, освещенности и др.) и сорта яблони, но прежде всего от температуры окружающей среды. Чем ниже температура, тем длиннее инкубационный период и наоборот. В естественных условиях он длиннее, чем, например, в оранжерее, где температура постоянная.
По данным различных ученых, инкубационный период в естественных условиях продолжается от 6 до 25 дней (табл. 2).
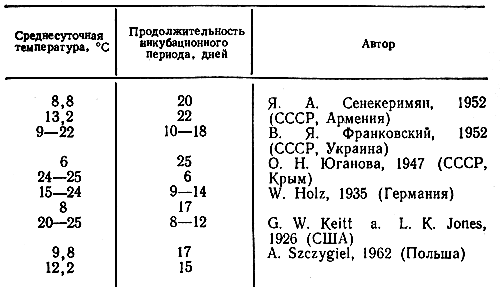
Таблица 2. Литературные данные о зависимости продолжительности инкубационного периода парши яблони от температуры при заражении аскоспорами гриба
Наши опыты по изучению длины инкубационного периода парши яблони велись с целью проверки литературных данных, а также получения собственных наблюдений в климатических условиях северо-западной зоны, так как в литературных источниках нет ни одного указания на опыты в сходных условиях. Так, О. Н. Юганова (1947) проводила свои опыты в Крыму, В. Я. Франковский (1952) — на Западной Украине, Я. А. Сенекеримян (1952) — в Армении.
Опыты проводились в садах учебно-опытного хозяйства ЛСХИ методом искусственного заражения листьев яблонь сильно поражаемого сорта Пепин-китайка в течение весны и лета 1966 г.
Еще задолго до распускания листьев в фазу зеленого конуса и до образования в перитециях па перезимовавших листьях зрелых аскоспор производилась изоляция подопытных веток яблони пергаментными мешочками. Первое заражение было проведено 20 мая при появлении первых листочков на ветвях яблони, последнее — 6 июня, так как в это время уже было отмечено появление конидиальной стадии гриба в естественных условиях.
В I декаде мая произошло массовое образование зрелых аскоспор гриба, которые могли вызвать первичную инфекцию яблонь.
Для приготовления инокулюма использовались многочисленные зрелые перитеции с перезимовавших листьев яблони. Суспензия с одинаковой для всех опытов концентрацией — 20 аскоспор в поле зрения микроскопа — наносилась каплями на листья изолированных веток. Ветки до инокуляции и после нее увлажнялись водой из пульверизатора, затем снова изолировались мешочками на 3 дня до первого просмотра изолятов. В дальнейшем просмотры велись ежедневно. Для каждого опыта суспензия готовилась заново с той же концентрацией опор.
Проведенные наблюдения свидетельствуют, что для прохождения инкубационного периода необходима определенная сумма температуро-часов, которая в среднем равна 4023,1 градусо-часа, что соответствует 167,6 градусо-дня.
При повышении температуры воздуха длина инкубационного периода уменьшается, при этом выявляется закономерность: при температурах, близких к оптимальным, увеличение на 1°С сокращает инкубационный период на один день. Так, при 12°С он равен 14 при 13°С - 13, при 14°С - 12, при 15°С — 11, при 16°С — 10 дням.
Зная количество градусо-дней, необходимое для прохождения инкубационного периода при заражении аскоспорами, можно вычислить длину этого периода в днях при различных среднесуточных температурах и тем самым проверить экспериментально полученные данные (табл. 3). В нашем опыте полученные данные оказались сходными с вычисленными, которые показывают, что вероятная длина инкубационного периода колеблется от 6,9 дня при температуре 24°С до 20,9 дня при 8°С.

Таблица 3. Сравнение вычисленных и экспериментально полученных данных инкубационного периода гриба
На основании полученных данных построена инкубационная кривая (Р. Н. Зленко, 1968), которая позволяет предвидеть срок появления конидиальной стадии гриба после первичного заражения яблонь аскоспорами гриба и сигнализировать защитные опрыскивания.
Таким образом, парша яблони в северо-западной зоне в своем развитии имеет следующие особенности: во-первых, источником первичной инфекции являются аскоспоры гриба, т. е. обязательным условием развития гриба является образование половой сумчатой стадии; во-вторых, сроки рассеивания аскоспор полностью зависят от местных метеоусловий и тесно с ними связаны, что обосновывает необходимость краткосрочных местных прогнозов заражений яблонь; в-третьих — аскоспоры гриба нашей местности имеют свои отличительные границы прорастания и оптимальные условия прорастания.
Особые метеорологические условия Северо-Запада накладывают свой отпечаток и на длительность инкубационного периода возбудителя парши яблони. Вышеприведенные биологические особенности гриба и особенности проявления болезни характерны и для других районов.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://berrylib.ru/ 'Садоводство и огородничество'